31.08.-05.09.26 Изумруд А.Непритворенный
01.07.-18.07.26. Чусовая. А.Чистяков
03.01.-08.01.27. Изумруд
"Цыплята С.Роуза".
Супер кратко об исследовании, которому С.Роуз посвятил более 20 лет жизни. Он изучал на примере цыплят, с помощью нейрофизиологических и нейрохимических методов, как они обучаются избегать определенных стимулов, и в каких именно областях мозга остаются следы (энграммы) о пережитом опыте. Роуз С. "Устройство памяти. От молекул к сознанию": Пер. с англ. - М.: Мир, 1995. - 384 с.Преамбула: любой объект обладает совокупностью свойств и качеств. Бусины, которые Роуз в ходе эксперимента добавлял в пшено для цыплят, имели с пшеном похожую форму и размеры, но были разного цвета (синие и красные, пшено желтое), и только бусины одного цвета он предварительно окунал в раствор с крайне неприятным вкусом. Что происходило дальше? Цыплята клевали пшено, иногда случайно клевали бусины не имевшие неприятного вкуса, например, синие. Поняв по вкусу, что это не пшено, цыплята выплевывали синюю бусину и продолжали клевать пшено. Но вот когда они случайно клевали красную бусину, имеющую крайне неприятный вкус, то они тоже выплевывали бусину и при этом трясли головой. Было видно, что они испытывают крайне неприятную эмоцию, в данном случае, отвращение. Что же изучал (хотел понять) Роуз в ходе своих экспериментов? Как возникают ассоциативные связи между различными свойствами разных модальностей? Как цыпленок обучается избегать бусин определенного цвета и вкуса. Какие изменения происходят в мозгу цыпленка? Вот это он и изучал двадцать с лишним лет.
Если вы поймете логику эксперимента, то вам станет понятна роль эмоций во всех случаях описанных на предыдущей странице (см. "Связывание нейронных сетей с позиций нейробиологии". Англосаксы это давно поняли, поэтому крайне эффективно формируют нужные им события по всему миру ( см. "Манипуляции. Дэйл Карнеги"), "Метод, по которому организуют гос.перевороты по всему миру".



















а) являются следствием опыта,
б) воспроизводятся, т. е. повторяются, у одного животного или у группы животных
в) имеют адаптивный характер.
Если не вводить критерий адаптивности, то образование рубцовой ткани тоже можно было бы считать формой памяти! С другой стороны, возможны обстоятельства, в которых животному нелегко найти оптимальную адаптивную стратегию, как, например, в слишком сложной ситуации или в случае, когда выбранная стратегия оказывается ошибочной. Однако и здесь нельзя говорить об отсутствии научения, что будет показано в этой главе несколько позднее. Поэтому смысл таких терминов, как «опыт» и «адаптивное изменение», достаточно сложен, иногда даже неоднозначен, и об этом никогда не следует забывать. Вспоминание — это проявление измененной поведенческой реакции спустя некоторое время после первоначального обучения.
Пластичность и специфичность.
Опыт — это термин из лексикона поведения. В переводе на язык биологии он означает пластичность. Для того чтобы эффективно функционировать, т. е. адекватно реагировать на окружающие условия, все живые организмы должны обладать двумя противоположными свойствами. Они должны сохранять стабильность (специфичность) в процессе развития и в зрелом возрасте, изо дня в день в течение всей жизни сопротивляясь непрерывному воздействию случайных изменений окружающей среды. Одновременно они должны быть пластичны, т. е. способны приспосабливаться и изменять свою специфичность по мере приобретения опыта.
Если некогда биологи говорили об организме как продукте взаимодействия природы и воспитания (на современном языке — генов и окружающей среды), то теперь такое противопоставление считают слишком упрощенным, поскольку основой как специфичности, так и пластичности служит проявление генов в процессе развития индивидуума. Без генов, обеспечивающих формирование мозга, способного обучаться на опыте, мы не смогли бы выжить. Однако мы не выжили бы и без генов, определяющих правильную систему связей в нашем мозгу в период развития. Объяснить диалектику специфичности и пластичности и понимание ее механизмов — одна из главных задач современной биологии. Изменения, происходящие в мозгу в результате накопления опыта, представляют собой форму пластичности, а память — одно из важнейших отображений этого опыта. Таким образом, для понимания механизмов пластичности (памяти) необходимо понять механизмы специфичности. Если бы мозг не сохранял в основном стабильность, не расшатываемую опытом, мы не смогли бы жить, что и подтверждают судьбы Фунеса и Шерешевского. Упорядоченные изменения имеют смысл только на относительно неизменном фоне.
Истоки поведения.
Адаптивное поведение как обязательное условие выживания свойственно всем организмам, однако многие процветающие формы жизни прекрасно обходятся без научения и вспоминания, даже без мозга. Возьмем подсолнечник, соцветие которого всегда обращено к источнику света, т. е. энергии. По мере роста подсолнух не учится на опыте, как лучше поворачиваться, и он не перестанет делать это, если после каждого поворота «наказывать» его электрическим ударом. Это поведение (поворачивание соцветия), носит оно адаптивный характер или нет, дано организму от природы, зафиксировано в его генетическом коде и программе развития. Такое присущее виду поведение (обычно говорят врожденное, но я избегаю пользоваться этим термином, так как он несет дополнительную идеологическую нагрузку) гарантирует адекватный ответ на специфические раздражители без научения методом проб и ошибок, Но за счет ограничения диапазона и гибкости реакций. Поведение изменяется не только у отдельной особи, но и в длинном ряду поколений как следствие эволюции. В отличие от этого приобретенные фирмы поведения вырабатываются на опыте, на практике; их преимущества — большая гибкость, свобода корректировки при изменениях окружающей среды и возможность учитывать наблюдаемые последствия при жизни данного индивидуума. Очень важно, однако, помнить, что приобретенные путем научения реакции не наследуются потомством. Передаются лишь степень пластичности и способность обучаться, которые в свою очередь «заданы» генетическим кодом и программой развития. Вот почему пора забыть наскучившие старые споры о врожденном и приобретенном. Чтобы познать мозг и поведение, нужно отрешиться от этого противопоставления и попытаться вместо этого понять диалектическую связь между специфичностью и пластичностью.
Первый критерий
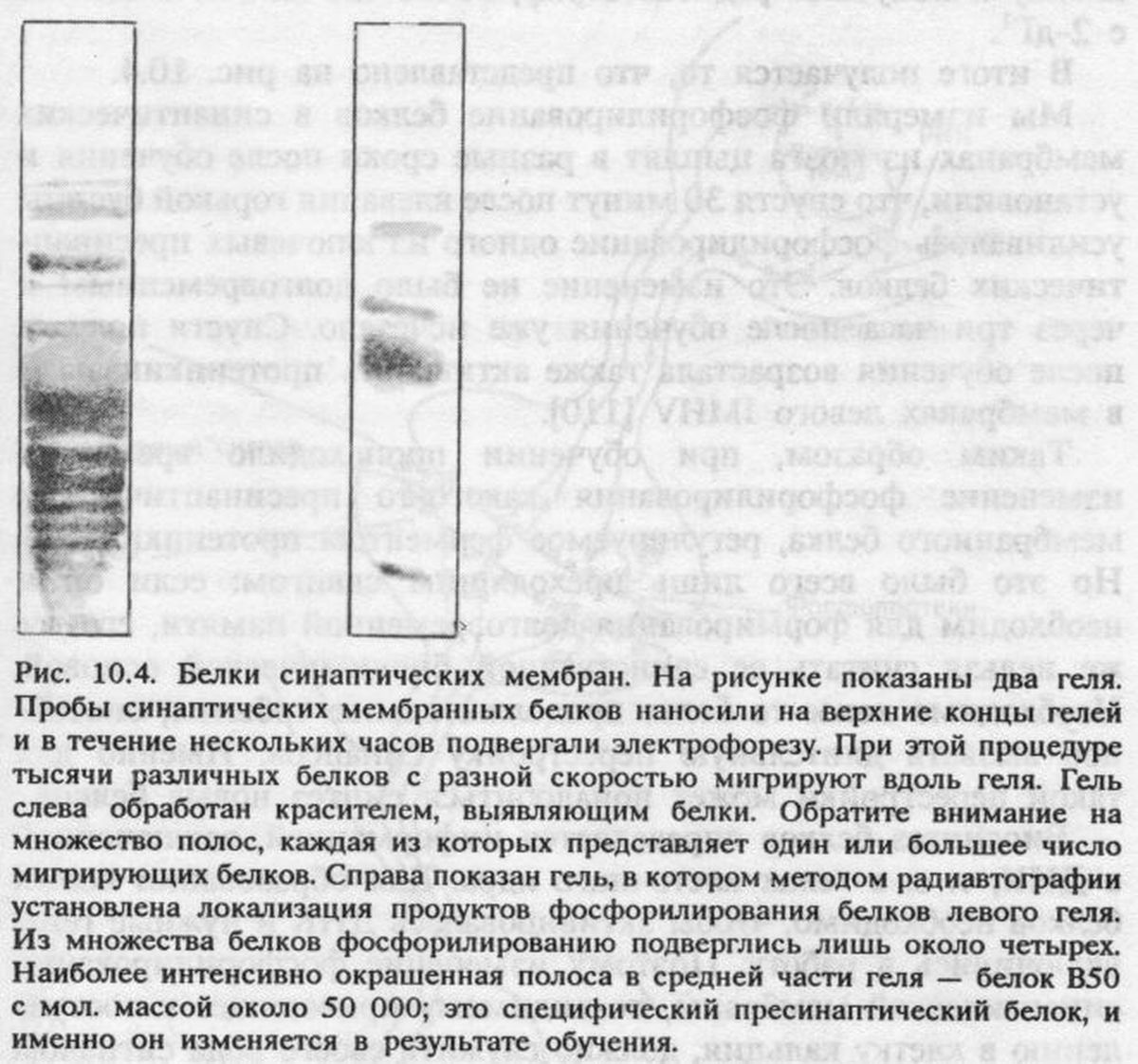
Предполагаемый фактор (система, вещество, скорость его образования или обновления) в локализованном участке мозга должен претерпевать количественное изменения в процессе формирования следов памяти. Если образуются новые или видоизменяются существующие синапсы, то в их химических или структурных компонентах должны появляться изменения, доступные для выявления биохимическими или микроскопическими методами (например, повышенное содержание синаптических мембранных белков, изменение размеров определенных синапсов или увеличение их числа). Однако если бы я выявил такое изменение в период обучения животного, но не убедился в его соответствии всем последующим критериям, я бы недалеко ушел от экспериментов шестидесятых годов, которые критиковал в предыдущей главе.
Второй критерий.
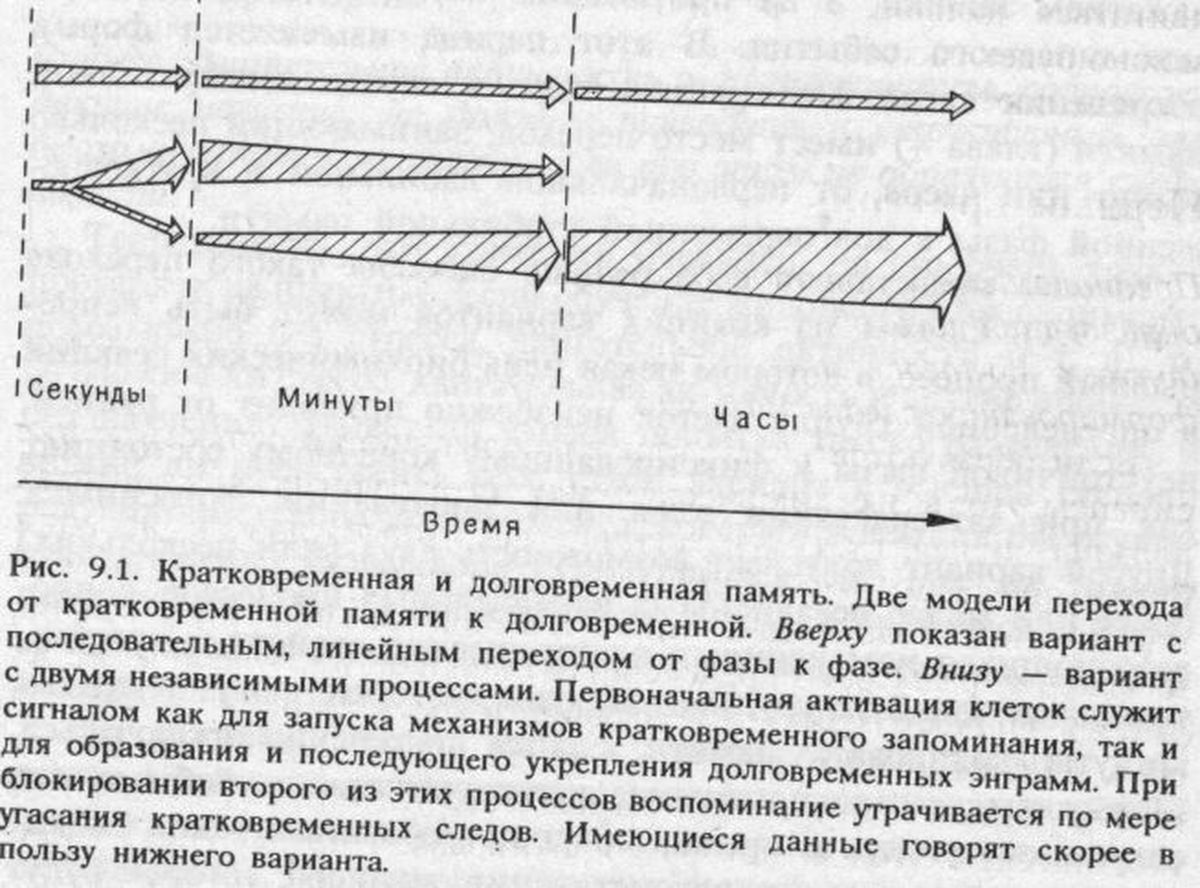
Изменение должно совпадать во времени с формированием следов памяти. Очевидно, что следы образуются не мгновенно, словно нажатием кнопки, а на протяжении нескольких часов после запоминаемого события. В этот период изменяется форма сохранения следа. По крайней мере, в случае декларативной памяти (глава 4) имеет место переход, занимающий несколько минут или часов, от первоначальной лабильной и кратковременной фазы к долговременной стабильной памяти.
Третий критерий.
Стресс, двигательная активность и другие процессы, сопровождающие научение, не должны приводить к структурным или биохимическим изменениям, если при этом не образуются следы памяти.
Четвертый критерий
Если клеточные или биохимические изменения ингибируются в период, когда должно было происходить формирование энграмм, то последнее блокируется и у животного возникает амнезия, и наоборот В этом случае изучаемый биохимический процесс, если он действительно специфически связан с образованием следов памяти, должен отсутствовать, когда отсутствует запоминание. Эксперименты, основанные на этом критерии, описаны в следующей главе.
Пятый критерий
Удаление участка, в котором происходят биохимические, клеточные или физиологические изменения, должно препятствовать образованию следов памяти (или вспоминанию) в зависимости от того, когда по отношению к времени тренировки произведено удаление.
Этот логический аналог двух предыдущих критериев тоже может казаться очевидным. Если изменения связей, формирующие след памяти, локализованы в отдельной небольшой группе клеток, а не распределены в мозгу диффузно, то удаление этих клеток приведет к исчезновению энграммы или помешает ее формированию. Поскольку в эксперименте довольно просто создать в мозгу небольшие локализованные «дырки», не вызывая общего поражения, нетрудно проверить справедливость утверждения, что какой-то участок является тем самым носителем энграммы, который определяет данное поведение. Если соответствующее повреждение приводит к амнезии или неспособности к обучению, это может подтвердить локализацию энграммы. Однако этот вывод отражает статичный и механистический взгляд на способ фиксации воспоминаний в мозгу. Если же процесс хранения следов более динамичен и в нем участвует много различных участков, такой эксперимент ничего не даст. Кроме того, игнорируется пластичность мозга (при удалении одного участка его функцию может принимать на себя другой участок); этот вопрос исследуется в экспериментах, описанных в главе 11. Наконец, никогда не следует забывать о свойственной всем травмирующим экспериментам двойственности: вспомним пример Ричарда Грегори с радиоприемником и подавляющим шумы транзистором.
Шестой критерий
При нейрофизиологической регистрации сигналов из мест изменения на клеточном уровне должны выявляться изменения в электрических ответах нейронов во время и (или) после формирования следов памяти.
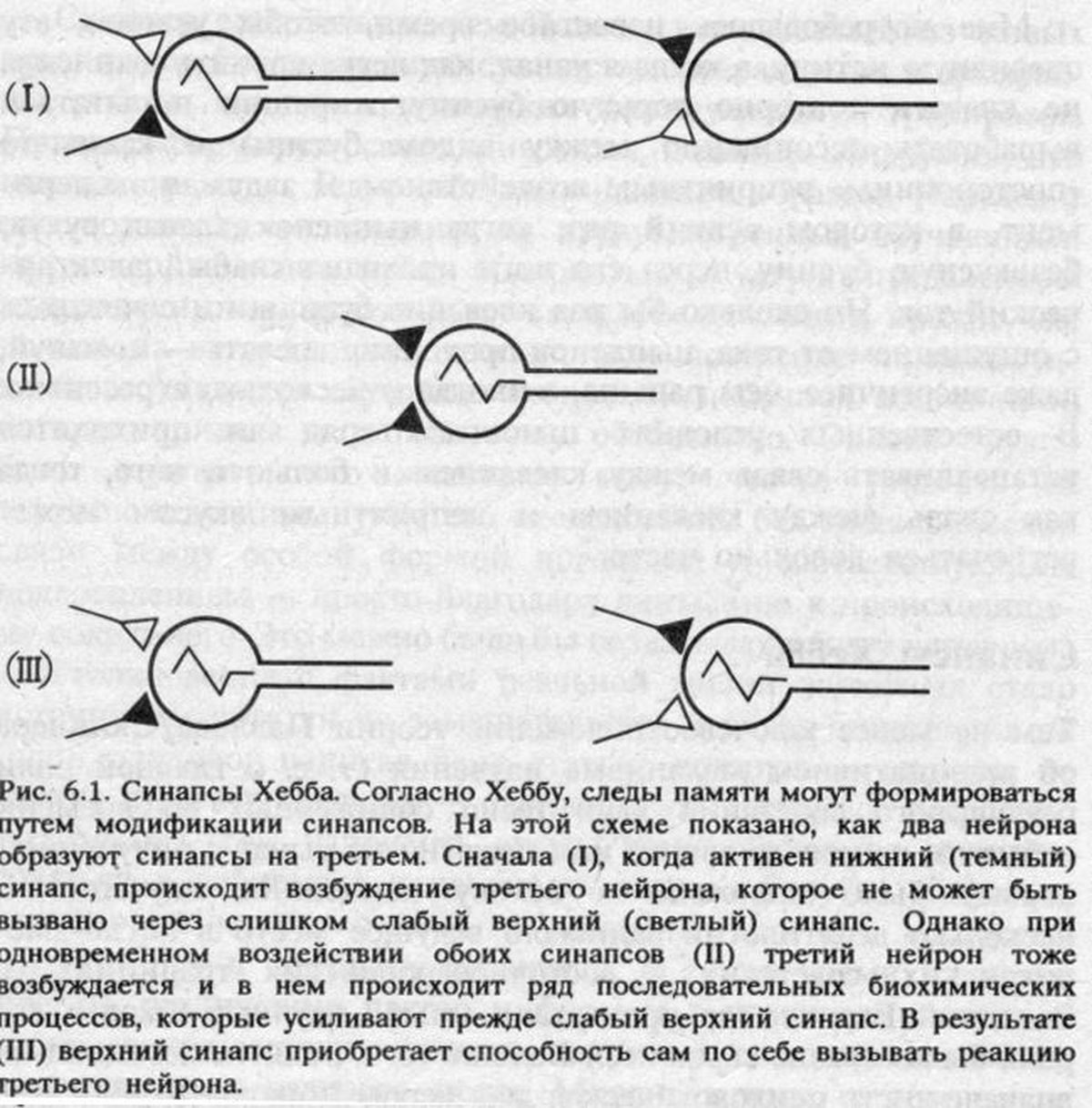
Если верна гипотеза Хебба о хранении следов памяти в форме измененных синаптических связей, то обучение должно сопровождаться изменением электрической активности нейронов, соединенных изменившимися синапсами.
Данная мною формулировка этого критерия предполагает, что начинать надо с поиска изменений на биохимическом и клеточном уровнях и уже на этой основе выявлять нейрофизиологические изменения, т. е. нейрофизиология в известном смысле рассматривается просто как побочный продукт биохимических и клеточных изменений.
Ключевым признаком ассоциативного научения (в отличие от привыкания и сенситизации) служит его долговременный характер, тогда как все рассмотренные до сих пор эффекты были кратковременными. Согласно моему второму критерию, при ассоциативном научении у аплизии должны возникать более стойкие клеточные изменения, соответствующие длительной модификации поведения. Хотя еще в начале 70-х годов было показано, что ингибиторы белкового синтеза не влияют на привыкание и сенситизацию, Кэндел только в середине 80-х годов переключил свое внимание на более долговременные клеточные процессы. Хотя введение этих ингибиторов не сказывается на привыкании, они вызывают амнезию в опытах с ассоциативным научением.
Гиппокамп как когнитивная карта
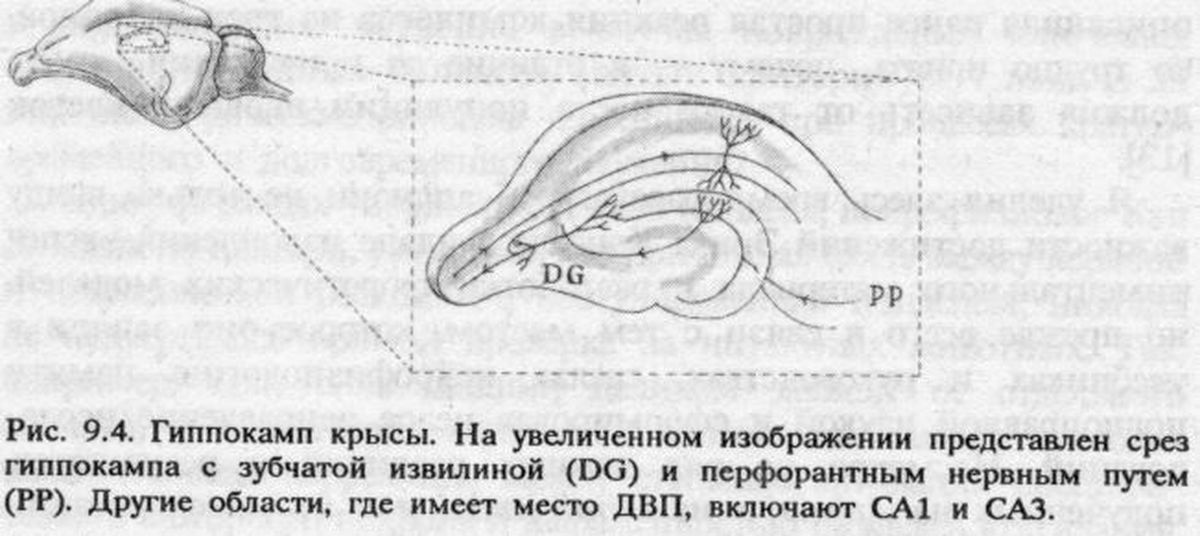
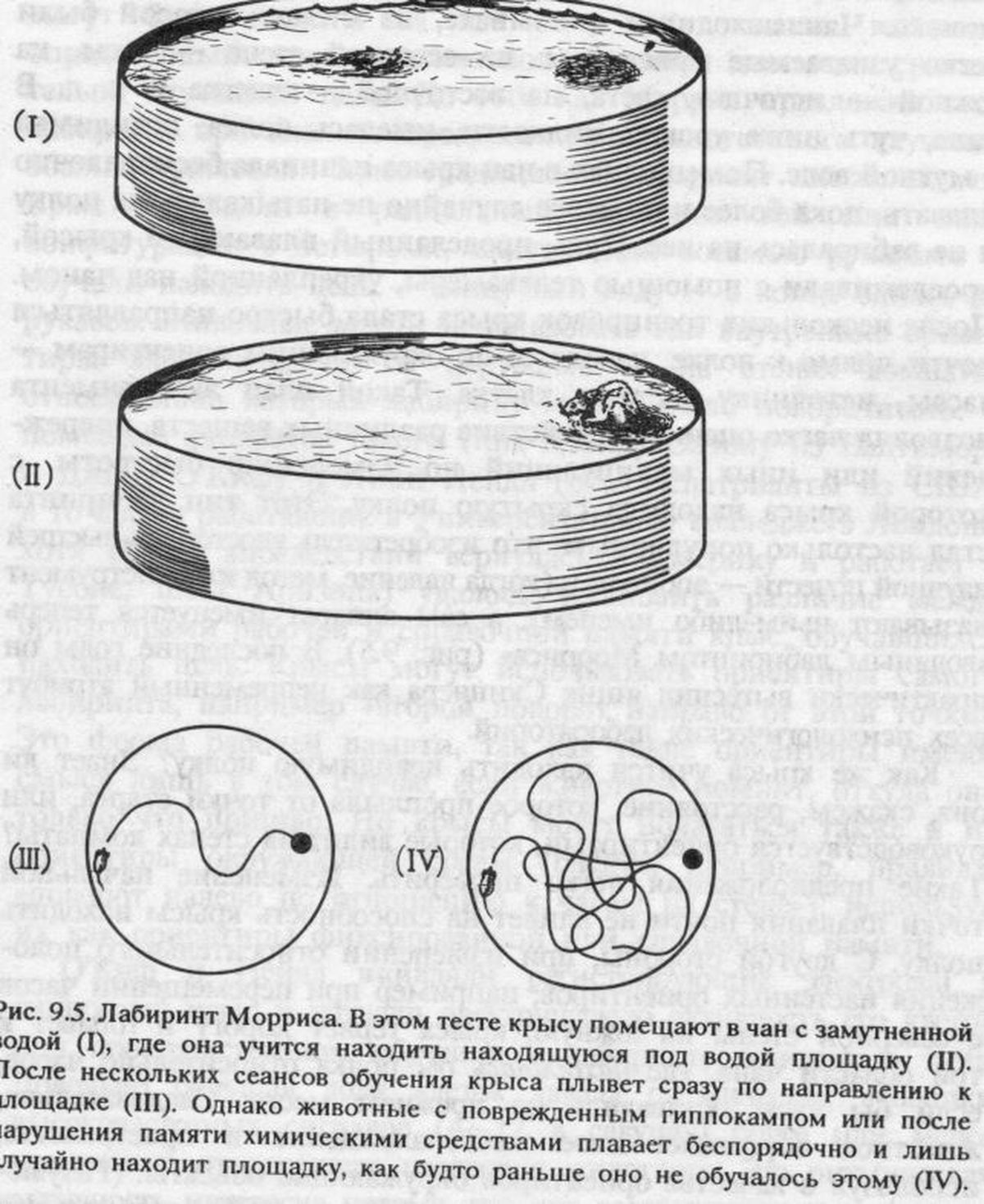
Признание ДВП в гиппокампе единственной моделью памяти в восьмидесятых годах скорее всего было закреплено все возраставшим потоком психологических данных о роли этого отдела мозга в обучении животных. В то время как исследования на людях позволяли предполагать участие гиппокампа в переходе от кратковременной декларативной памяти к долговременной (что подтверждалось наблюдениями над обезьянами с поврежденным гиппокампом), опыты на крысах выявили еще один аспект его роли в механизмах памяти. Одним из наиболее выраженных последствий повреждения гиппокампа у этих животных была утрата способности обучаться задачам на пространственную ориентацию, например в лабиринтах.
Итак, Черкин вместе с Илен Ли-Тенг натолкнулся на мысль предлагать цыплятам клевать маленькие разноцветные бусины. Если бусина имела горький вкус, так как исследователи смочили ее спиртом, хинином или едким метилантранилатом, цыплята, однажды клюнув ее, выражали отвращение, энергично тряся головой и вытирая клюв о пол клетки, а когда через несколько секунд или суток им снова предлагали сухую бусину такого же цвета, они всякий раз пятились назад и отказывались клевать. Такова была в своей основе модель разового обучения пассивному избеганию, которой я заинтересовался.
Обучение было разовым, так как цыпленку достаточно было клюнуть бусину однажды, чтобы извлечь урок. Это была выработка реакции избегания, поскольку птенец переставал делать то, что делал бы в иных условиях. Это было пассивное избегание, так как цыпленок должен был лишь воздерживаться от контакта, не предпринимая никаких активных действий, чтобы избежать неприятного ощущения.
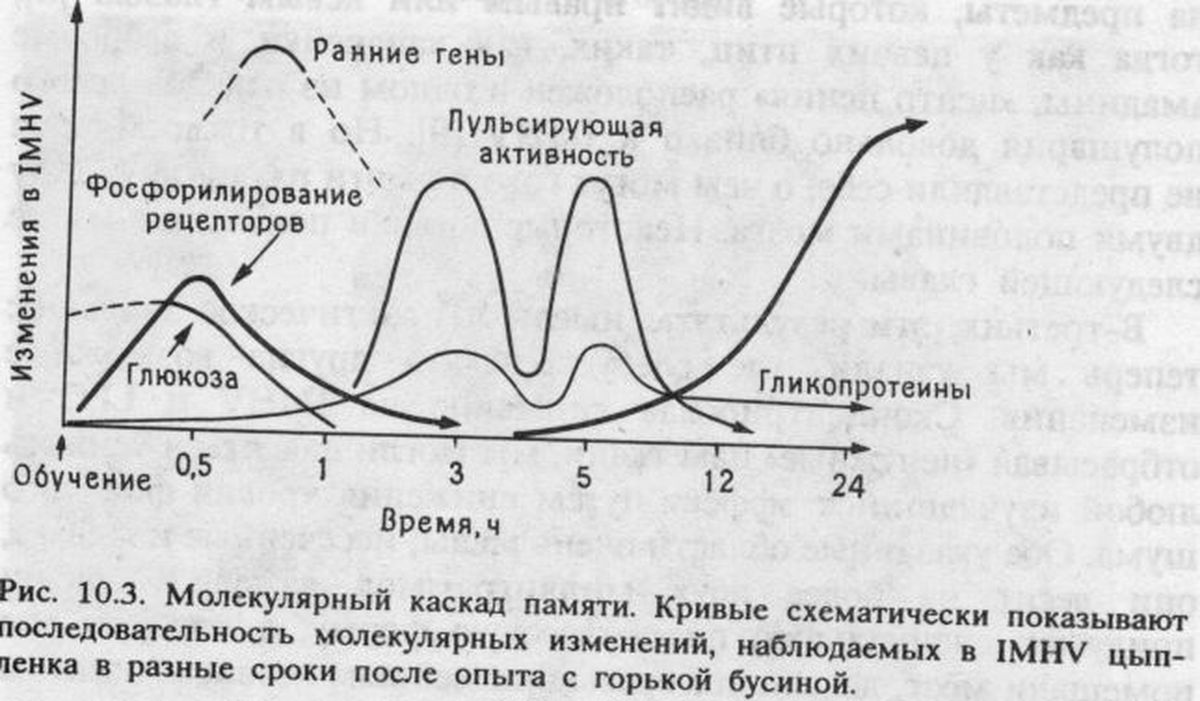
Через 30 минут после того, как цыплята клевали горькую бусину, содержание мускариновых рецепторов возрастало в той же самой области мозга, где мы наблюдали изменения при импринтинге. Но это повышение носило временный характер и сохранялось не более трех часов после обучения. Это, по-видимому, означало, что увеличение количества мускариновых рецепторов было связано с ранними фазами формирования следов памяти.
Конечно, этот вывод нельзя было считать окончательным, что сразу становилось ясно при обращении к моим критериям, изложенным в предыдущей главе. Например, наблюдавшееся увеличение могло быть реакцией на вкус самого метилантранилата, а не следствием сформировавшейся ассоциации между клеванием бусины и ее вкусом.
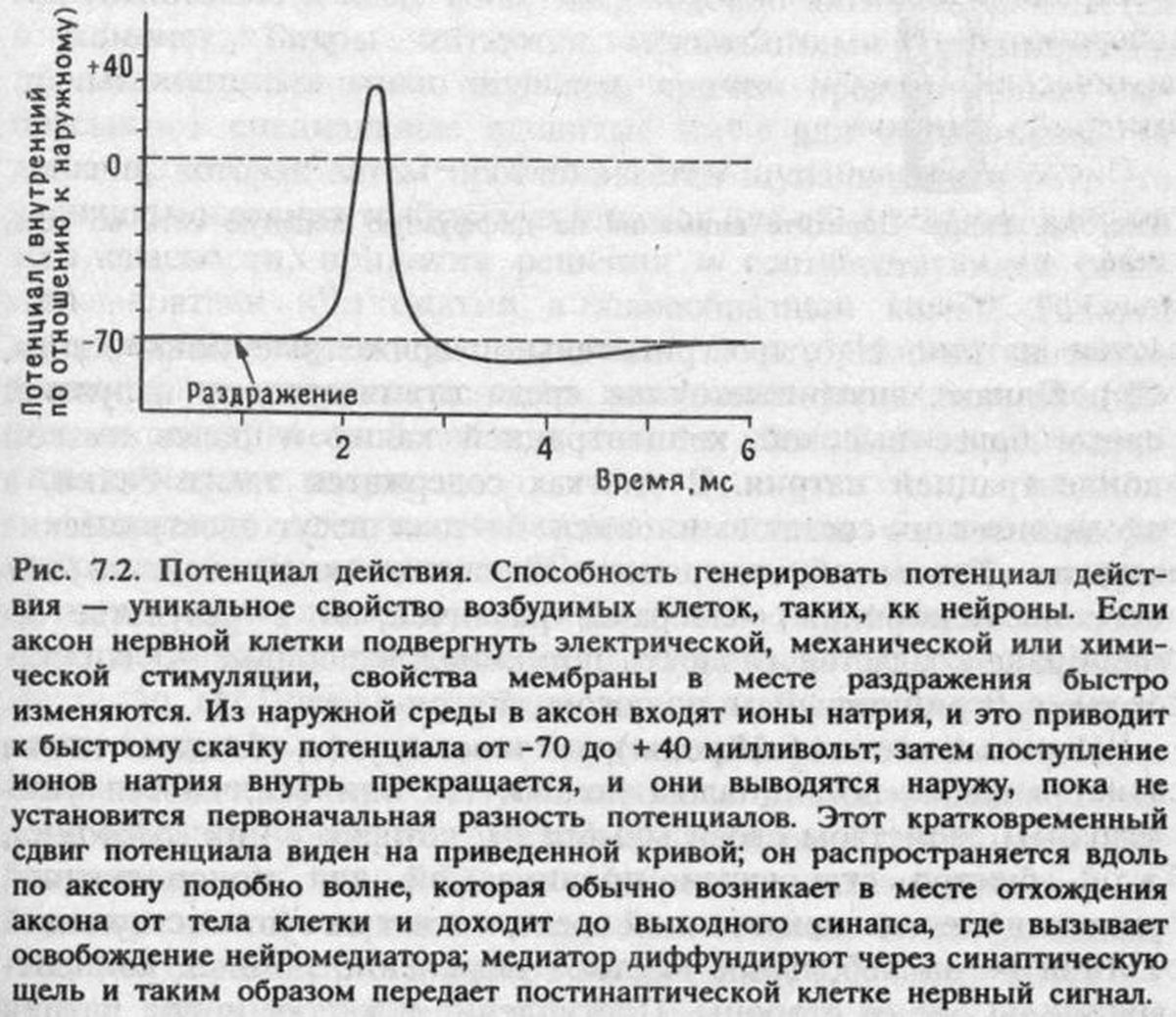
Размышляя над решением этой проблемы, я натолкнулся на статью Ларри Беновица, нейробиолога из Гарвардского университета, опубликованную в начале семидесятых годов. В ней описывалось, как цыплят обучали пассивному избеганию и сразу после этого пропускали через голову слабый электрический разряд, что, по-видимому, заставляло их забыть связь между видом и вкусом бусины: при повторном предъявлении они клевали ее так же энергично, как и в первый раз. Но если ток пропускали не ранее десяти минут после обучения, память о приобретенном опыте сохранялась и цыплята избегали повторно клевать бусину [16]. Это можно было объяснить тем, что ранние этапы образования энграмм связаны с электрической активностью нейронов, а она нарушалась при воздействии тока сразу после обучения; при более позднем пропускании тока эти этапы успевали завершиться и процесс мог беспрепятственно идти дальше.
Таким образом я смог проверить данные Беновица. Он оказался прав: если ток пропускали уже через минуту после обучения, у большинства цыплят развивалась амнезия, а при задержке на десять минут цыплята избегали клевать бусину повторно.
Четвертый критерий: приводит ли подавление биохимических процессов к подавлению памяти?
Логика экспериментов с ингибированием самоочевидна, но на протяжении ряда лет я отказывался от их проведения, так как мне оставалось не ясно, что конкретно может дать применение ингибиторов с широким спектром действия, например ингибиторов белкового синтеза, для познания биохимических процессов, которые я пытался расшифровать. Но когда мы подошли к более детальному изучению отдельных звеньев биохимического каскада, я убедился, что использование достаточно специфических ингибиторов может пролить свет на молекулярные механизмы. Например, мы обнаружили, что если перед началом обучения вводить вещества, блокирующие долговременную потенциацию в гиппокампе и пространственное научение (ингибиторы рецепторов глутамата NMDA-типа), то у цыплят развивается амнезия. К таким же последствиям приводит инъекция в левое полушарие протеинкиназы перед самым обучением или сразу после него [20].
Таким образом, эксперименты как с электрошоком, так и с ингибиторами показали, что для образования следов памяти необходим биосинтез специфических гликопротеинов [21].
Итак, для того чтобы в мозгу цыпленка сформировалась ассоциативная связь между клеванием бусины и горьким вкусом, приводящая к стойкому изменению поведенческой реакции, в определенной области переднего мозга должен произойти ряд последовательных событий. Эти события завершаются структурной модификацией синапсов и дендритов, и в итоге проявляются также в изменении электрических свойств клеток, в частности в характере их ритмической активности на протяжении нескольких часов после обучения. Все это, видимо, удовлетворяет шестому критерию.
Подобно всем научным данным, мои данные на самом деле не что иное, как результаты регистрации измерений, записи на бумажных лентах, цифры на шкале или экране приборов (философ-позитивист и физик Эрнст Мах в начале века назвал такие наблюдения «показателями стрелок», я манипулирую ими, пытаясь понять их значение, а поняв, произвожу обратную экстраполяцию, чтобы сделать выводы о поведении молекул, клеток и организмов в реальном мире.
Мы спланировали эксперименты так, чтобы один человек производил повреждение мозга, второй обучал цыплят, не зная о характере повреждения, а третий (обычно это был я) проводил последующие испытания, опять-таки не зная о сделанном двумя другими.
Сомнений быть не могло: цыплята с поврежденным IМНV явно обучались пассивному избеганию, т. е. клевали горькую хромированную бусину, так же, как их ложно оперированные собратья, трясли головами, ощутив ее вкус, и отворачивались при повторном предъявлении. Но спустя несколько часов при вторичном испытании они полностью забывали приобретенный опыт и клевали сухую бусину столь же энергично, как и контрольные цыплята, которым раньше давали бусину, смоченную водой. Таким образом, повреждение мозга не влияло на поведение птенцов (клевание бусины), на чувство вкуса или общую подвижность — они лишь не помнили, что следует избегать бусин определенного вида.
Итак, первый парадокс: для запоминания необходим интактный левый IМНV, но коль скоро цыплята уже усвоили задачу (по крайней мере в первый час после обучения), IМНV оказывался ненужным [2].
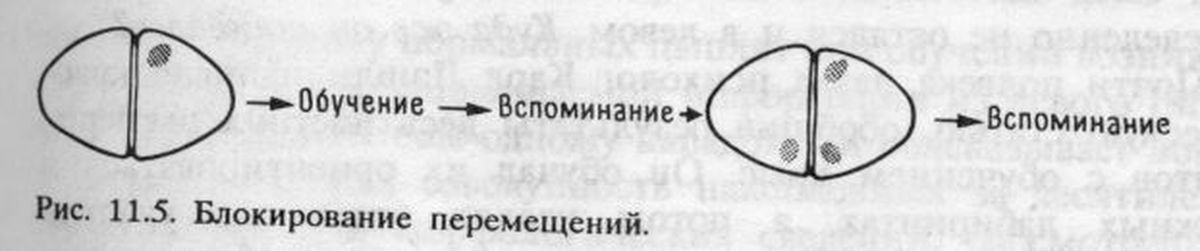
Куда же переместилась память? Учитывая все полученные ранее биохимические и морфологические данные, следовало искать ее следы в LРО. Эксперимент подтвердил это. Двустороннее повреждение LРО через час после обучения действительно вызывало амнезию, которой не было при одностороннем правом или левом повреждении. По-видимому, в норме след памяти после обучения каким-то путем мигрирует из IМНV в LРО, чем и объясняется отсутствие амнестического эффекта повреждения после тренировки. Это был весьма интересный результат, так как он позволял понять, почему мы находим биохимические и морфологические изменения и в IМНV, и в LРО. Тот факт, что для сохранения памяти достаточно одного LРО, правого или левого (одностороннее повреждение не вызывало амнезии!), согласовался с тем, что многие из выявленных нами изменений имели место как в правом, так и в левом LРО, т. е. след памяти о бусине и реакции избегания сохранялся в обоих полушариях (рис. 11.2).
Если пассивное избегание вырабатывается и при отсутствии LРО, то не происходит ли реорганизации следа памяти каким-то иным образом? Может быть, в этом случае след просто закрепляется в IМНV? Если это так, то повреждение LРО до тренировки и повреждение IМНV после тренировки — две операции, по отдельности не приводящие к амнезии, — при их сочетании у одного и того же цыпленка должны будут вызвать амнезию. Так оно и оказалось в действительности (рис. 11.3). Получив такие результаты, я предположил, что решающая роль принадлежит IМНV. Но Дейв думал иначе. Тогда мы произвели односторонние повреждения, и выяснилось, что он прав. У цыплят, не имевших LРО во время обучения, последующее повреждение левого IМНV не влияло на запоминание, а повреждение правого IМНV вызывало амнезию. Все это схематически показано на рис. 11.3.
Результаты были интригующими. Размышляя над ними, я решил, что все можно объединить, исходя из несколько механистической модели, в которой первичный след памяти о бусине и реакции избегания образуется в левом IМНV, а затем, спустя несколько часов после обучения, «перекочевывает» сначала в правый IМНV, а потом в правый и левый LРО (рис. 11.4).
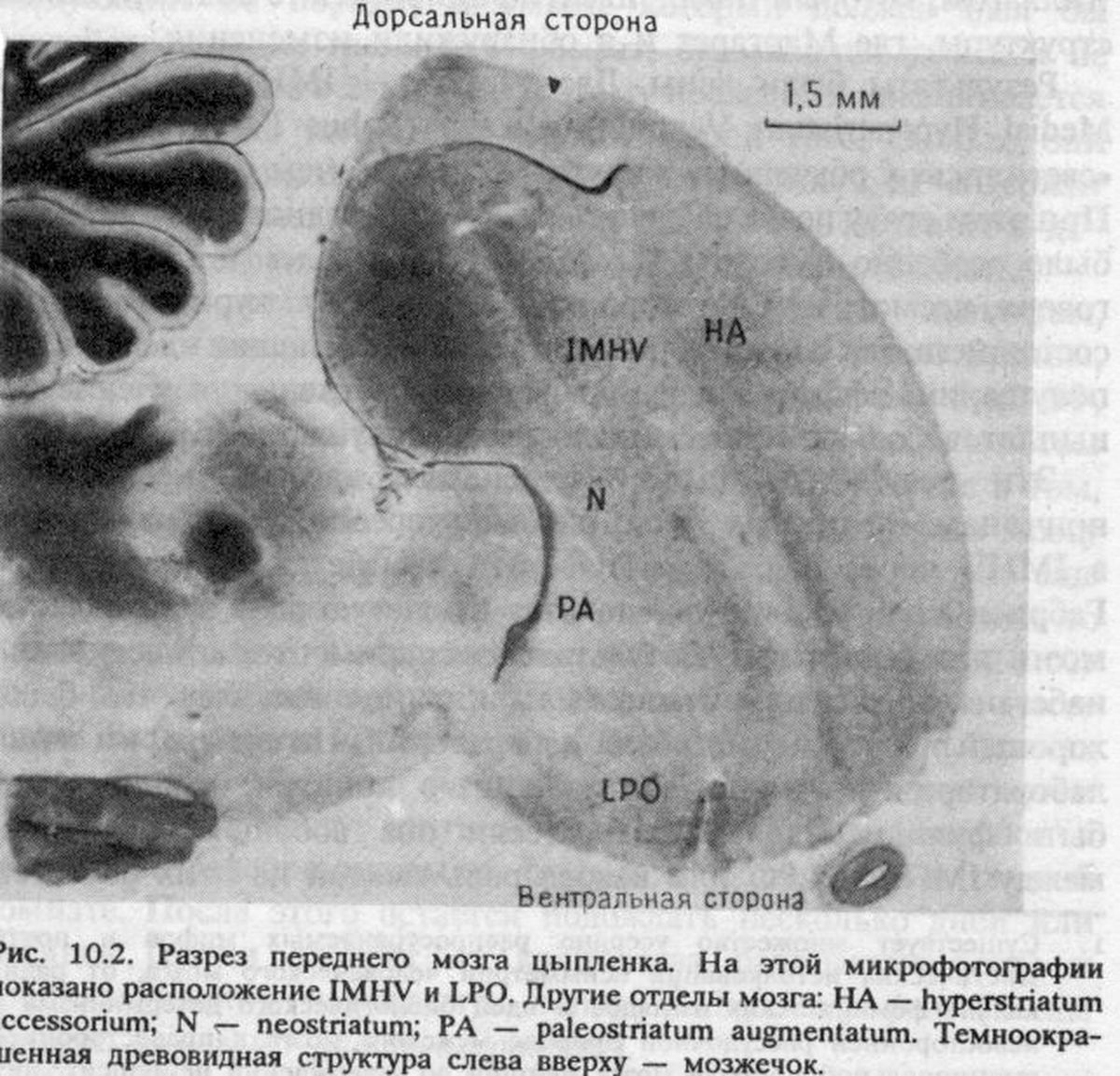
Я пришел к этой схеме исключительно на основании логики экспериментов с повреждением мозга. Но подобная схема шокирует нейроанатомов, у которых тут же возникает очевидный вопрос: разве есть какие-то прямые нервные пути между IМНV и LPO! Согласно простой концепции коннекционизма, такие пути действительно необходимы. В то время, когда мы приступали к исследованию, мы не знали, существуют ли они, но это казалось маловероятным. Нам было известно, что все нервные пути от органов чувств, таких как глаза и вкусовые рецепторы, сходятся к IМНV. Мы знали даже о непрямых связях между IМНV и LРО. Именно поэтому IМНV рассматривались как некие «ассоциативные зоны», где интегрируется разнообразная входная информация — например, сопоставляется зрительное восприятие бусины с ощущением ее вкуса.
До этого я был готов, как некий научный Пуаро, собрать всех подозреваемых в гостиной и указать виновного, как было совершено преступление, то бишь как работает память. Теперь это было исключено. Эксперимент дал однозначный результат — целиком отрицательный. Ничего, пустота, нуль, никакой амнезии. След памяти не мог перейти в правый IМНV, но он определенно не остался и в левом. Куда же он подевался?
Почти полвека назад психолог Карл Лэшли написал классическую статью, обобщив результаты десятилетних экспериментов с обучением крыс. Он обучал их ориентироваться в сложных лабиринтах, а потом удалял отдельные участки мозговой коры, чтобы выяснить, где сохраняются следы памяти. К своему удивлению, он не обнаружил специфических участков, удаление которых приводило бы к полной утрате усвоенного навыка; вместо этого навык постепенно ухудшался по мере увеличения размеров удаленной области коры. Создавалось впечатление, что память зависит просто от количества мозгового вещества. На основе этих наблюдений Лэшли построил концепцию «эквипотенциальности» коры. Свою статью он озаглавил «В поисках энграммы» и закончил ее выводом, что память одновременно присутствует в мозгу везде и нигде [4].
Опыты Лэшли и его неутешительные выводы утратили актуальность в результате последующих экспериментов, однако парадокс локализации памяти остался. Это напоминает мне, во-первых, о том, что при повреждении IМНV и LРО мы изучаем не функции этих областей, а функции остального мозга, которые быстро перестраиваются в отсутствие двух первых; и, во-вторых, о том, что память нельзя понимать как нечто статичное, «находящееся» строго в одном месте или в небольшой группе клеток; она существует в более динамичной и рассеянной форме. Кроме того, мозг — это тонко и сложно организованная структура с многообразными средствами функционального обеспечения. Вы мешаете ему осуществлять какую-то деятельность, а он находит другие пути к той же цели. Блокируете оптимальные связи между IМНV и LРО, а цыпленок находит им замену. Мозг действует не как набор стандартно соединенных мелких ячеек, а как функциональная система, насыщенная разнообразными связями. Конечно, всегда следует помнить об анатомии мозга, но нельзя замыкаться в ней, поскольку ясно, что и сам мозг не сводится к анатомически выявляемым структурам. Настаивать на его механистическом объяснении, игнорируя мои предостережения о неразумности редукционизма, — это значит обрекать эксперимент на погружение в трясину парадоксальных результатов.
Порядок из хаоса?
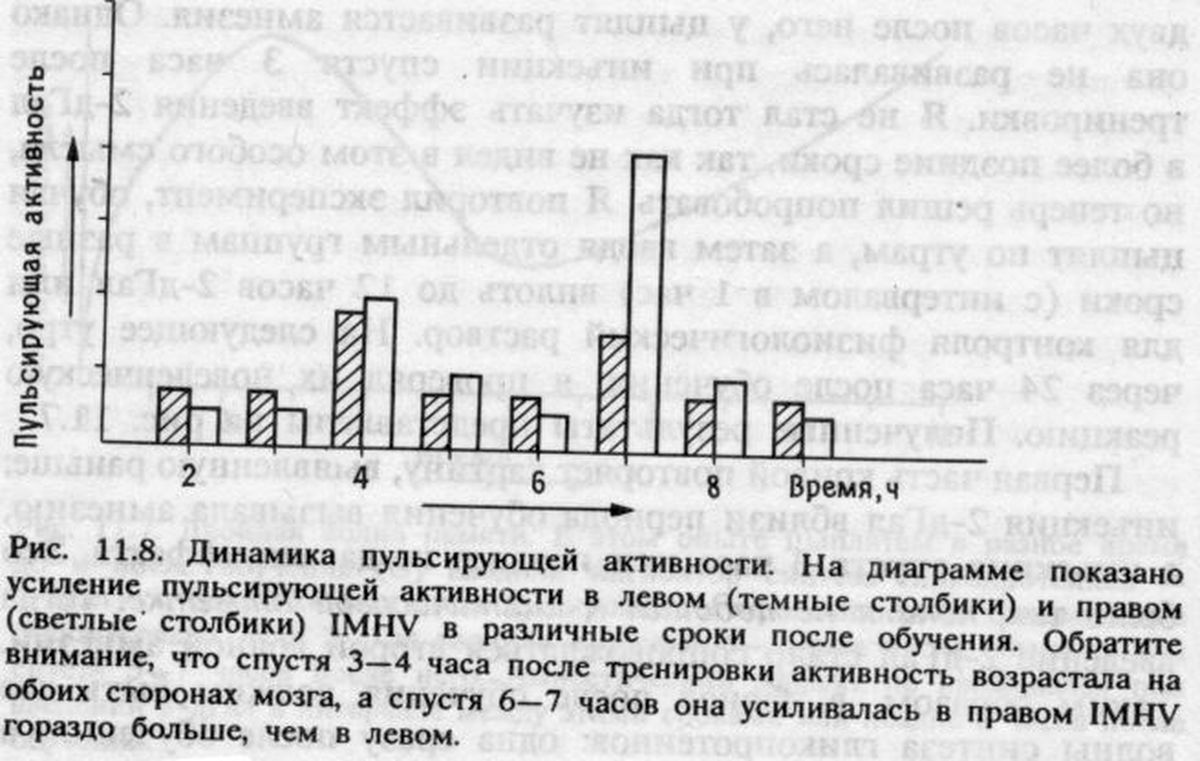
Когда вы участвуете в какой-нибудь исследовательской программе, нередко случается, что, проснувшись в понедельник утром с идеей новых экспериментов, вы мчитесь в лабораторию, чтобы тут же начать их; но проходят недели или месяцы, а у вас нет ничего, кроме неоднозначных результатов, которые никак не вписываются в общую картину, уже начавшую складываться в вашем воображении. Вы решаете прекратить опыты или опубликовать их в каком-нибудь малоизвестном журнале, надеясь, что никто не заметит, как классическое совершенство вашей теории превратилось в абстракционистский сумбур Джексона Поллока. Нечто в этом роде годами испытывали все мы в лаборатории. Но вдруг мне стало ясно, как согласовать мои теории с обнаруженной ранее двухфазной клеточной активностью. Если цыплятам вместо бусины, смоченной очень горьким метилантранилатом, предложить что-нибудь с менее резким вкусом, например ту же бусину, смоченную очень слабым раствором метилантранилата или хинина, у них тоже появится реакция избегания и она будет сохраняться несколько часов, после чего они забудут о горьком вкусе бусины. Предположим, что эта «слабая» память связана только с первой волной клеточной активности в IМНV, которая затем не передается в форме более стабильной второй волны в LРО: такие слабые воспоминания индуцируют только первую, но не вторую волну биохимических изменений. Оказывается, дело обстоит именно так [5]. Более того, в экспериментах с выработкой «условнорефлекторного отвращения к пище», описанных в главе 6, было установлено, что даже акт клевания сухой бусины вызывает первую волну биосинтеза гликопротеинов.
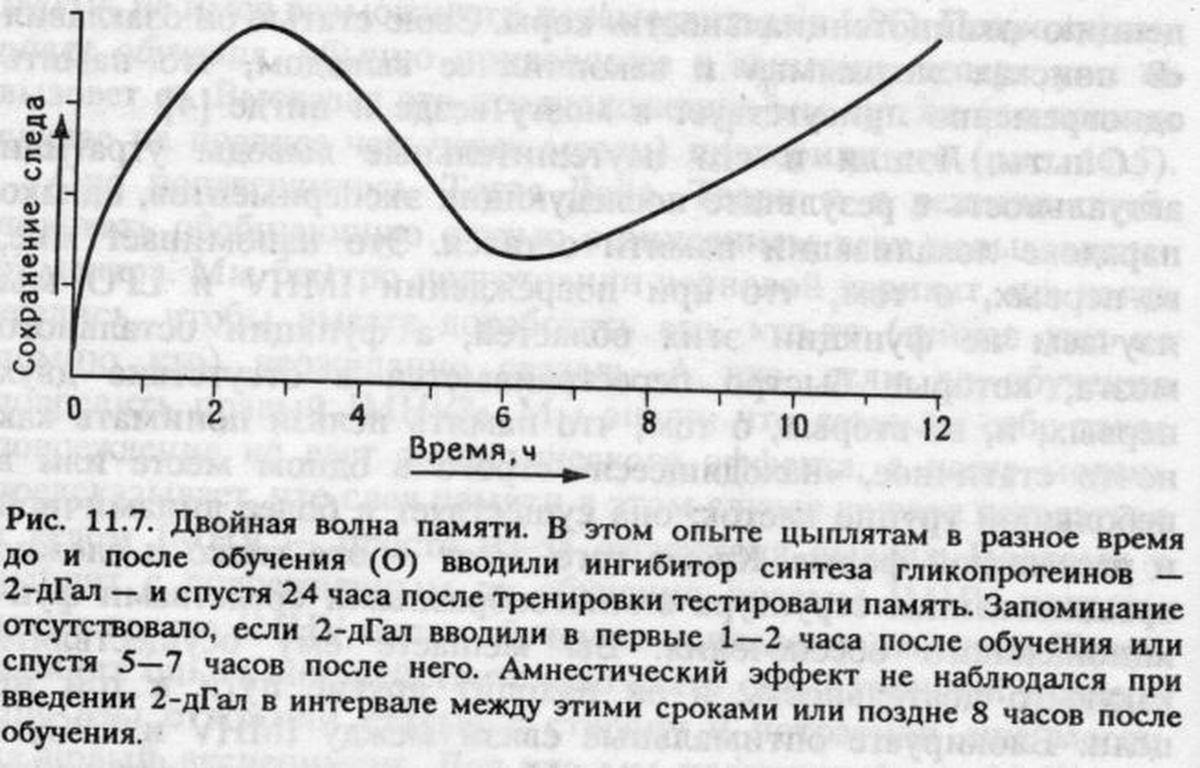
Дело в том, что цыпленок все время наблюдает, замечает, исследует и запоминает различные факторы окружающей его действительности, хотя бы потому, что поначалу еще не знает, что важно помнить, - у него формируется нечто вроде эйдетической памяти, как у ребенка. В обедненных условиях клетки с ее монотонным интерьером появление даже вовсе безвкусной бусины вносит новизну, которую цыпленок по меньшей мере не может не заметить, он клюет бусину и память о ней запечатлевается, скорее всего в IMHV. Если потом, как в эксперименте с условным избеганием, цыпленок чувствует легкое недомогание, то он приходит к вполне разумному, хотя и ошибочному, выводу, что причина этого – именно клевание бусины. В очень простом эксперименте с подавлением синтеза гликопротеинов 2-D-галактозой мы показали, что для образования такого следа необходима первая волна их биосинтеза (рис. 11.9) [6].
Пока все хорошо. Теперь я могу свести воедино картину процессов кратковременной и долговременной памяти, локализованных в разных отделах мозга, объединенных потоком информации между ними и связанных с двумя волнами клеточной активности. Однако я еще не выбрался на гладкую дорогу. Долговременная память оставляет в левом IMHV длительно сохраняющийся след, но после того, как воспоминание сформировалось, этот IMHV цыпленку больше не нужен, чтобы помнить о горькой бусине. Как это можно понять?
Я не знал этого. Когда я начал писать книгу, то собирался закончить эту главу парадоксом превращения порядка в хаос. В первоначальном варианте она оканчивалась следующими словами: Этим рассказом я и хотел бы закончить главу. Понимайте его как хотите: с одной стороны, это серия строгих экспериментов для выявления необходимых и достаточных клеточных процессов в определенных областях мозга, которые в целом создают след простой ассоциативной памяти, изменяющий последующее поведение цыпленка; с другой стороны — параллельное описание памяти как ускользающего от наблюдения динамического процесса, столь важного для цыплят, что его не удается окончательно связать с определенными местами и «выключить».
Теперь это не годится. Это никого не устроит: ни меня, ни издателя, ни вас, моих читателей. Конечно, я мог бы сделать нечто большее. В начале этого года, совершенно неожиданно, решение стало так ослепительно ясно, что мне оставалось только изумиться, как туго моя мысль была затянута в смирительную рубаху механистического редукционизма. Все это время я рассказывал, как цыплята клевали хромированную бусину, будто она была для них простым, неразложимым объектом. Ибо мы сами считали ее такой — простой хромированной бусиной, и думали, что так же считают цыплята. Но, конечно, есть множество причин, по которым этого не должно быть. Клевавший горькую бусину цыпленок не имел понятия о том, что в ней важнее: цвет, величина или форма, время дня или место, где она появлялась. Не зная, на чем остановиться, цыпленок, для того чтобы выжить, должен был учитывать все эти особенности. Он не мог сразу же сделать вывод, что важна только бусина данной конфигурации. Поэтому цыпленку необходимо было классифицировать то, что он видел и испытывал.
Предположим теперь, что из всех свойств бусины важны цвет, размеры и форма (есть основания думать, что это действительно так, но я не буду углубляться здесь в подробности [7]). После того как цыпленок познакомился с бусиной, любой из этих признаков в дальнейшем помогает избегать ее. Это очень похоже на попытку вспомнить забытое имя: мы вызываем в воображении лицо человека, подбираем имена, которые, как нам кажется, близки по звучанию к забытому, или последовательно пробуем все буквы алфавита. Все это — разные стратегии поиска признаков забытого имени.
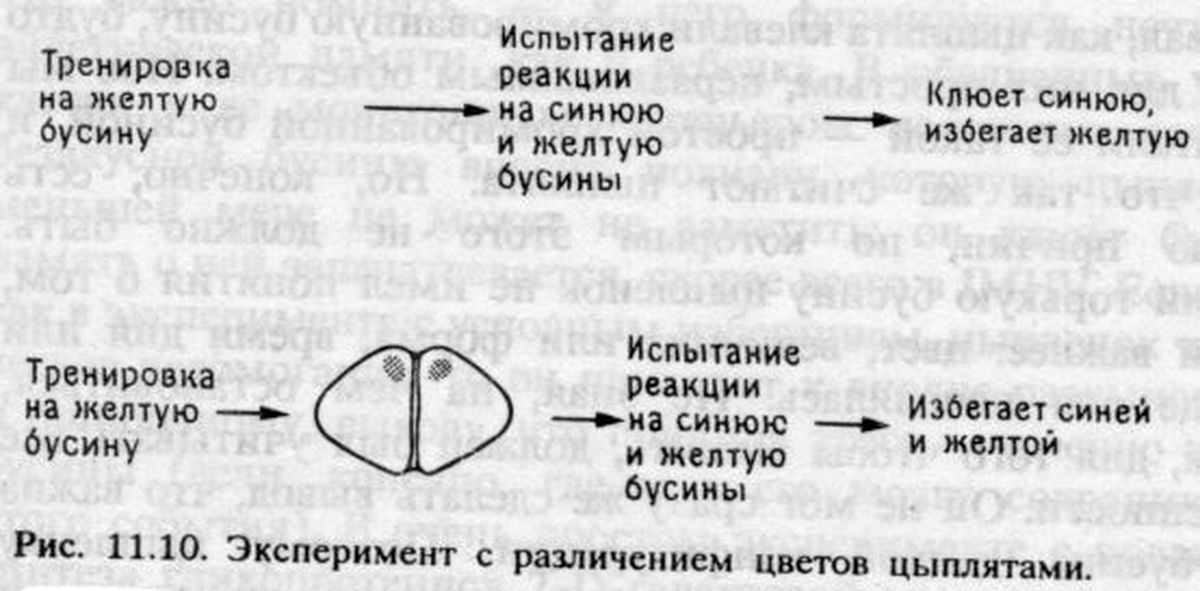
Если цыпленок действительно помнит не «бусину», а набор ее ключевых признаков, то последние не обязательно должны «храниться» в одном и том же участке мозга. Возможно, информация о цвете бусины сохраняется в одном месте, о форме — в другом и т. д. Просто мы никогда не пытались проверить такое предположение, а вместо этого обучали цыплят не клевать хромированную бусину и потом наблюдали их поведение, повторно предъявляя такую же бусину. Но если информация о цвете хранится в IMHV, а о других признаках в LРО, становится понятно, почему после обучения цыплятам не нужен IМНV, чтобы вспомнить бусину: в их LРО отложилась информация о других ее особенностях. В то же время в IМНV долгое время сохраняются клеточные изменения, так как «цветовое представительство» бусины все еще существует в этом отделе. Таким образом, мы нашли способ избежать парадокса Лэшли.
После того как я обдумал все это, проверка не составляла особой трудности. Нужно было лишь обучить цыплят не клевать горькую желтую бусину, а потом предложить им на выбор две бусины: ту же самую и новую — синюю. Цыплята с неповрежденным мозгом будут клевать вторую и отвергнут первую. А как поступят цыплята с повреждениями? Если память о цвете хранится в IМНV, удаление LРО до обучения ни на чем не скажется: птенцы будут по-прежнему обучаться и помнить различие между бусинами, как и в предшествующих экспериментах (см. рис. 11.2). Но теперь удаление IМНV будет иметь последствия: хотя у цыплят сохранится способность к научению, в последующих тестах они будут избегать обеих бусин — и желтой, и синей, так как без IМНV у них останется информация только о величине и форме бусины, находящаяся в LPO, по этим признакам они не смогут отличать одну бусину от другой и будут избегать всех мелких предметов округлой формы, подозревая, что они горьки на вкус.
К лету 1991 года мы с Терри уже провели такой эксперимент [8]., тем самым был найден выход из редукционистской ловушки и, признав мозг открытой обучающейся системой, мы воссоздали порядок из хаоса.
Послесловие от руководителя проекта GO-RA: Возможно, этот материал поможет вам в понимании того, как именно формируется наше мировоззрение, как работает наше сознание и подсознание, а также заставит внимательнее отнестить ко всем материалам с пометкой в названии "Связывание". И еще один важный момент: наш мозг подозрительно похож на аналоговый компьютер, у которого меняется передаточная функция при изменении нейронных структур. Просто элементная база другая нежели в обычных компьютерах.
Важно: изменения в поведении цыпленка, в том что у него появилось отвращение к бусинкам определенного цвета и формы, явилось следствием того, что у этих бусинок был крайне неприятный вкус. Это в свою очередь означает, что цыпленок испытывал в этот момент большие эмоции.
Это связывание привело к изменению нейронных структур, и эти изменения не зависели от желания цыпленка - это происходило автоматически.
О подсознании и сознании. "Субличности".Что такое свойства, качества, навыки, умения. Подробнее:
Эти книги заслуживают того, чтобы прочитать их целиком:
Ильюченок Р.Ю., Эмоции и память. СО АН СССР, Новосибирск, 1988.
Роуз С. Устройство памяти. От молекул к сознанию: Пер. с англ. - М.: Мир, 1995. - 384 с.
Дойдж Н., Пластичность мозга. М., Эксмо, 2010
Поделиться: